Connective Tissue on a Divergent Spectrum Part Two: The Nose, Mouth & Fascia
Explore the impact of collagen genomic variation across the senses
CONNECTIVE TISSUEBIOLOGY
Alexandra Chambers
12/26/20254 min read
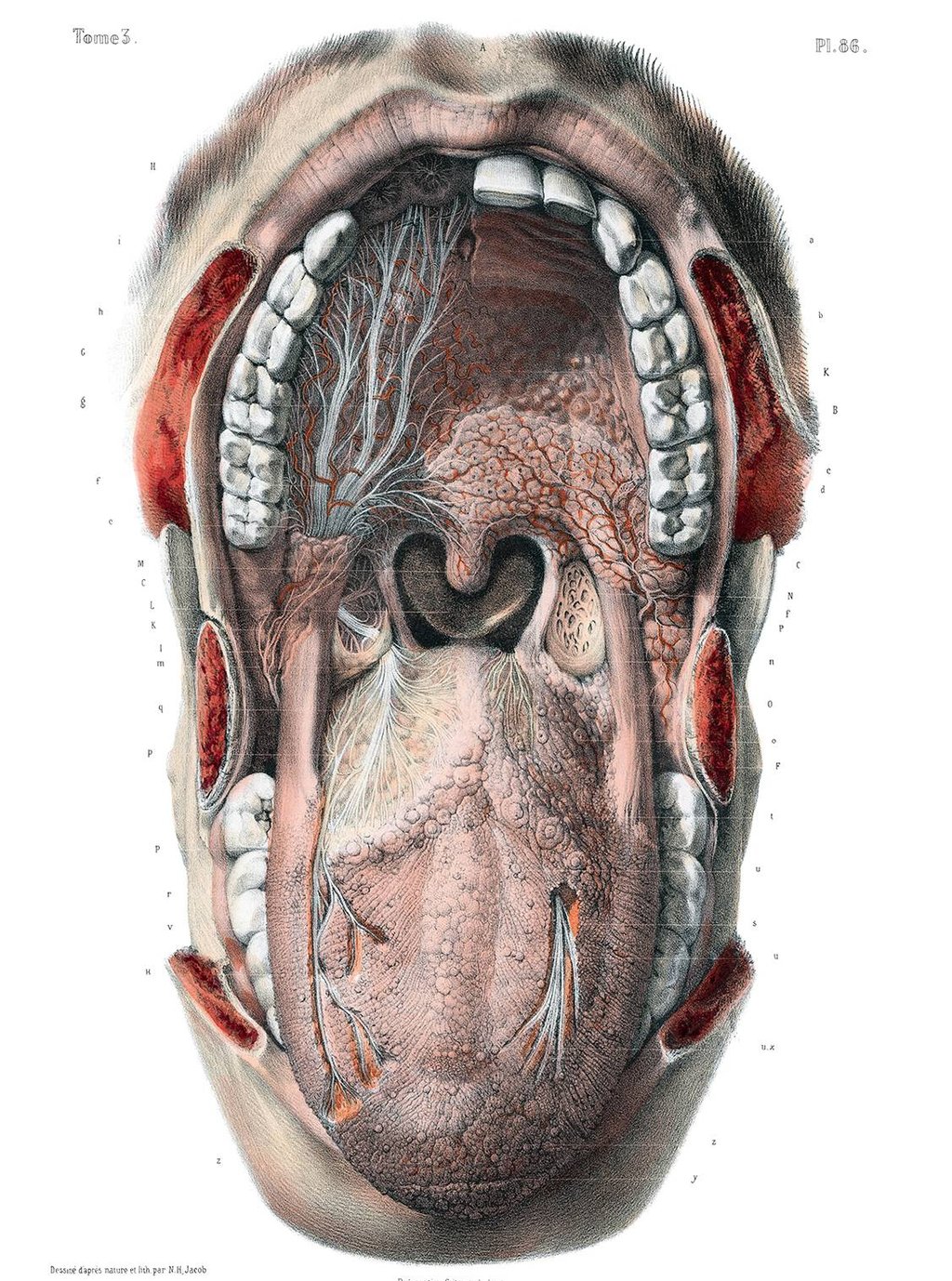
While sensory sensitivities are often framed as behavioural, neurological, or psychological, very few models account for the tissue-based architecture underlying those sensations. Connective tissue plays a structural and bioelectrical role in shaping perception across all sensory domains. Nowhere is this more visible (and less understood) than in the face.
Nose
The olfactory system is deeply integrated with both fascia and the autonomic nervous system. The connective tissue surrounding the olfactory bulb and cribriform plate (a thin, sieve-like bone at the top of the nasal cavity) can influence the transport of molecules and cerebrospinal fluid. This may explain why some individuals have heightened or distorted smell perception. Inflammation, laxity, or subtle cranial structural differences - common in people with hypermobile-type connective tissue divergence - can alter olfactory signalling.
Olfactory receptors live in a moist, mucus-lined epithelium deep within the nose. To smell something, molecules must dissolve into this layer and bind to receptors - which are embedded in a structural scaffold of tissue and supported by the extracellular matrix (ECM). That’s connective tissue.
If that matrix is too permeable, fragile, or inflamed, it alters the sensory landscape completely: smell becomes muted, distorted, or hypersensitive. Many divergent individuals report extreme reactions to smell - not just an aversion, but overwhelm. This may reflect altered signal strength at the very root of olfactory detection. In addition, olfactory neurons are directly exposed to the external environment, bypassing the blood-brain barrier (BBB). This makes them vulnerable to environmental toxins, pathogens, and oxidative stress - a vulnerability that may be amplified in individuals with less robust connective tissue scaffolding.
Mouth
The architecture of the mouth is often overlooked in sensory and developmental assessments. The lingual frenulum (the connective band under the tongue) varies considerably between individuals. On one end of the connective tissue spectrum is tongue-tie (ankyloglossia), where the frenulum is short or tight, restricting tongue movement. On the other end is an often-overlooked phenomenon seen in some individuals with Ehlers-Danlos Syndrome (EDS) or related connective tissue divergence: a lax or even absent frenulum. This can also be accompanied by soft oral tissue, poor muscle tone, or delayed oral-motor coordination.
Other midline variants - like bifid uvula, submucous cleft palate, or high-arched palate - are frequently overlooked, yet they often co-occur in collagen-related divergence and may subtly affect speech, feeding, and sensory integration.
These variations reflect broader shifts in tissue integrity and coordination, with downstream effects on feeding, speech, taste, and breath control. The hard palate and soft palate can also show signs of structural variance - high-arched or soft/drooping palates being common among neurodivergent populations. This may affect nasal resonance, chewing, or even breathing during sleep.
Taste receptors are housed in mucosal tissues embedded in a collagenous matrix. Inflammatory signalling, mast cell activation, or altered tissue perfusion can all affect taste perception - something often missed in clinical or behavioural framings of food aversion.
Fascia
The skin is the body’s largest sensory organ, and its responsiveness is shaped by more than just nerve endings. The extracellular matrix, including collagen and elastin, plays a role in how mechanical stimuli (like pressure, texture, temperature) are received and transmitted. People with altered collagen expression may experience touch as painful, itchy, irritating, or even electrically charged.
Connective tissue also plays a major role in proprioception (body awareness), as fascia houses sensory receptors that help regulate movement and positioning. Loose or dysregulated fascia can contribute to sensory processing challenges, or hyper-awareness of internal sensations.
Mechanosensitive ion channels like Piezo1 and Piezo2, embedded within connective tissue scaffolds, play key roles in translating pressure into neural signals - meaning that connective tissue tone directly modulates how we feel the world.
As with neurodivergence itself, connective tissue variation presents across a non-linear spectrum. Not everyone will have every trait - and some will show only subtle signs that would likely go unrecognised by typical diagnostic thresholds.
A new language is needed for these patterns - one that dissolves artificial divides between neurology, anatomy, and perception. A language that recognises connective tissue as both sensor and signal; a terrain in which biology, ancestry, and environment co-produce experience. Without this shift, we will continue misreading sensory divergence as pathology rather than divergent and varied physiology.
Connective tissue is more than a structural element; it is a major sensory component.
Some collagen/connective-tissue variants may represent evolutionary trade-offs. These may be emerging from developmental architectures that favour greater neural complexity, altered sensory processing, or expanded cranial-neural patterning, even if those same architectures may reduce resilience elsewhere in the body.
Altering the developmental matrix can alter the physical architecture of the developing brain.
There is now substantial evidence that joint hypermobility, HSD and EDS occur disproportionately alongside autism, ADHD and other neurodevelopmental presentations. A Swedish population study found substantially elevated recorded diagnoses of autism and ADHD among people diagnosed with EDS, while sibling findings suggested that shared genetic or early developmental factors could contribute.
Some people with collagen/connective tissue genomic variants may have high cognitive potential or distinctive cognitive strengths. However, this potential becomes obscured when the body is chronically burdened by broadly hostile environmental conditions. The same structural vulnerability that affects tissue integrity may also increase susceptibility to systemic overload, pain syndromes, dysautonomia, fatigue, and brain fog, causing their true baseline capacities to be underestimated or missed entirely.
Are collagen-divergent people being systematically mischaracterised because research measures them on their presentation after environmental injury, chronic stress, and multisystem burden have suppressed/altered their baseline cognitive and physiological potential?
Is fibromyalgia, in at least a substantial subset, better understood as an environmentally burdened state occurring in structurally and metabolically vulnerable bodies, rather than some primary standalone syndrome?
As neural architecture becomes more complex, natural selection may be favouring changes in extracellular matrix and connective-tissue organisation that support cranial development, cortical expansion, sensory complexity, and neural patterning - but some of those same shifts may have carried trade-offs in whole-body mechanical robustness.
It may be that a more complex brain structure requires a different developmental matrix environment - different tissue mechanics, different cranial growth dynamics, different ECM signalling, different balance between flexibility and structural rigidity.
Therefore,
collagen / ECM differences are not faults, as such
they may instead reflect participation in a different developmental architecture
that architecture could support increased neural complexity or sensory-cognitive divergence
but at a cost of reduced tolerance for brute-force biomechanics, repetitive load, and risk of injury due to the increasing exposome/biochemical environment (chiari/ cervical cranial instability/ spontaneous cerebrospinal fluid (CSF) leaks/ functional neurological disorders (FND).
Progressive research should ask what developmental architecture is present before the exposome accumulates - and what might that neurological architecture be capable of under different environmental conditions today?
Artist: Nicolas-Henri Jacob, for Jean-Baptiste Marc Bourgery, 1831-54
Contact
Reach out with questions or collaboration ideas.
AChambers@divergentgenomics.org
© Alexandra Chambers 2026. All rights reserved.