Connective Tissue on a Divergent Spectrum Part One: The Ear and Eye
Explore the impact of collagen genomic variation across the senses
CONNECTIVE TISSUEBIOLOGY
Alexandra Chambers
12/26/202510 min read
Sensory overload in divergent populations is frequently conceptualised as a consequence of altered central sensory processing, attentional filtering, perceptual intolerance, or behavioural regulation. Within autism, ADHD, sensory processing difference, migraine, hyperacusis, auditory processing disorder, and related presentations, sensitivity to light and sound is often interpreted primarily through neurological or psychological models. Although these frameworks may describe part of the downstream response, they do not fully account for the biological structures through which sensory information first enters the body.
This article proposes that visual and auditory sensitivity should also be examined as embodied sensory-interface phenomena. Before light or sound is interpreted by the nervous system, it must pass through specialised tissues whose optical, mechanical, pressure-regulatory, and extracellular matrix properties shape the quality of incoming sensory information. The eye and ear are connective tissue-rich organs whose function depends on collagen, elastin, fibrillin, extracellular matrix organisation, tissue compliance, fluid dynamics, and regulated mechanical tension.
A connective tissue framework does not replace neurological models of sensory processing. Rather, it situates them within a broader biological pathway. If the peripheral sensory organs are structurally divergent, the signal arriving at the nervous system may already be altered. Sensory overload may therefore arise not only from cortical interpretation, but from multi-level interactions between connective tissue genomics, extracellular matrix architecture, epigenetic regulation, inflammatory repair, fibrotic remodelling, altered pressure transmission, neural gain, and downstream processing demand.
Connective tissue variation is often recognised only when it meets the threshold for a named syndrome, such as Ehlers-Danlos syndrome, Marfan syndrome, Stickler syndrome, or another diagnosed connective tissue disorder. This approach is insufficient as connective tissue expression exists across a non-linear spectrum, ranging from overt syndromic presentations to subtler forms of tissue fragility, altered compliance, pressure sensitivity, or matrix instability that may never be formally diagnosed.
There are individual presentations with recognisable systemic features, including hypermobility, translucent skin, mitral valve prolapse, craniofacial differences, vascular fragility, lens instability, scoliosis, or skeletal involvement. There are also individuals whose connective tissue differences may present less visibly, but still influence daily function. These may include light sensitivity, migraine, visual fatigue, ear pressure, sound intolerance, tinnitus, dizziness, gastrointestinal dysmotility, chronic fatigue, postural symptoms, pain, or poor tolerance of complex sensory environments.
Such variation may be influenced by genomic divergence in collagen, fibrillin, elastin, extracellular matrix, methylation, redox, inflammatory, immune, and tissue repair pathways. Epigenetic regulation during development and across the lifespan may further modify expression. Environmental exposures, infection, inflammation, oxidative stress, mechanical strain, injury, and chronic tissue remodelling may also shape the extent to which connective tissue vulnerability becomes functionally apparent. Introgressed ancestral variants, including Neanderthal-derived variants affecting immune and tissue architecture, may represent an additional layer of biological diversity relevant to divergent physiology.
This framing has particular relevance for divergent populations because sensory sensitivity is often interpreted at the level of outward response rather than biological origin. What is described as sensory sensitivity may, in some individuals, reflect a material difference in the tissues that receive, filter, transmit, and regulate sensory input.
The Eye
The eye is commonly discussed as an extension of the nervous system, but it is also a collagen-rich optical organ. Its ability to receive and regulate light depends on the structural integrity of the cornea, sclera, iris, lens capsule, zonular fibres, retina, Bruch’s membrane, and associated vascular and extracellular matrix systems. These tissues do not merely support vision; they shape the optical signal before it is converted into neural information.
The cornea and sclera are heavily dependent on collagen types I, III, and V. Their organisation influences transparency, curvature, tensile strength, and optical stability. The iris contributes to light regulation through tissue structure and pigment distribution. Zonular fibres, which suspend and stabilise the lens, are rich in fibrillin-containing microfibrils and are therefore relevant to Marfan-type spectrums and other fibrillin-related tissue presentations. The lens capsule is collagen-structured and vulnerable to oxidative and mechanical change. Bruch’s membrane and the retinal extracellular matrix contribute to retinal support, nutrient exchange, and tissue resilience.
Where these structures are thinner, looser, stiffer, irregular, oxidatively stressed, mechanically unstable, or less able to buffer light effectively, visual input may be altered before cortical interpretation begins. Light may scatter unpredictably. Contrast sensitivity may be reduced. Accommodation may require greater effort. Retinal signalling may become more easily overloaded. Glare, flicker, contrast, and visually dense environments may impose a higher physiological burden.
For the individual, this may present as photophobia, migraine, visual snow, glare intolerance, eye strain, screen intolerance, motion sensitivity, visual fatigue, or difficulty functioning in bright or visually complex environments. These experiences are often interpreted as anxiety, avoidance, poor tolerance, poor attention, or behavioural resistance. However, such interpretations may describe the visible response while failing to account for altered optical mechanics and connective tissue architecture at the sensory interface.
Visual overload should therefore be considered as a whole-system event involving connective tissue, optical structure, retinal signalling, vascular regulation, oxidative stress, neurological processing, and environmental load. In this model, the visual system is not simply receiving too much information because the brain is failing to filter it. The visual signal itself may be formed under altered structural conditions.
The Ear
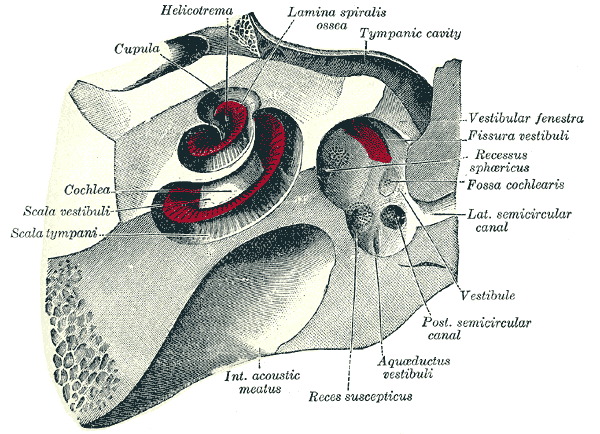
Hearing begins as a mechanical process. Sound enters the ear as vibration and is shaped by the tympanic membrane, ossicular chain, middle-ear ligaments, cochlear fluids, tectorial membrane, basilar membrane, vestibular structures, and surrounding bony and connective tissue boundaries before it is converted into neural signal. The auditory system is therefore deeply dependent on tissue mechanics.
The tympanic membrane contains organised collagen layers that regulate vibration. The ossicles and middle-ear ligaments depend on appropriate collagen and elastin tension. The tensor tympani and stapedius muscles modulate sound pressure and are anchored within connective tissue structures. The cochlea depends on precisely tuned stiffness, fluid movement, and matrix organisation. The basilar and tectorial membranes influence auditory amplification, frequency selectivity, and the transfer of vibration to sensory hair cells. The vestibular labyrinth relies on collagen-structured compartments and regulated fluid pressure for balance and spatial orientation.
Collagen-related auditory phenotypes are already recognised in conditions involving genes such as COL11A1, COL11A2, and COL2A1, where altered collagen architecture affects structures including the tectorial and basilar membranes and changes cochlear function. In COL11A2-related auditory phenotypes, disrupted organisation of collagen fibrils in the tectorial membrane has been linked to altered viscoelastic properties. This indicates that the hearing apparatus can be mechanically different at source, before any question of central auditory processing arises.
This genomic layer is significant because the ear does not only detect sound. It conducts, filters, dampens, amplifies, separates, and transforms vibration. If the connective tissue framework differs, the signal entering the nervous system may also differ. The result may be an increase or decrease in hearing, or potentially an altered quality of hearing: sharper frequencies, intrusive small sounds, poor damping of layered noise, abnormal resonance, distorted loudness growth, sound-triggered discomfort, or rapid auditory fatigue.
Epigenetic Regulation
The structure and function of sensory organs are not determined by DNA sequence alone. Inner-ear development, auditory function, tissue repair, vulnerability, and regeneration are also regulated by epigenetic mechanisms, including DNA methylation, histone modification, chromatin remodelling, and non-coding RNAs such as microRNAs. These mechanisms influence how genes are expressed in response to developmental timing, environmental exposure, inflammation, oxidative stress, noise exposure, injury, and ageing.
Epigenetics therefore belongs within the same framework as connective tissue genomics. A person may inherit variants affecting collagen, matrix organisation, redox balance, methylation, or repair capacity, but the functional expression of those variants may be modified across the lifespan. Noise exposure, infection, inflammatory signalling, mitochondrial stress, nutritional status, endocrine factors, and tissue injury may all alter the resilience of sensory structures.
This is also particularly relevant in divergent populations, where immune, metabolic, connective tissue, neurological, and sensory differences frequently overlap. A connective tissue variant may interact with epigenetic regulation, inflammatory history, oxidative burden, mechanical strain, and environmental exposure to produce a variable sensory presentation. The same person may also move between states of relative tolerance and sensory collapse depending on tissue load, inflammation, fatigue, hormonal state, or repair capacity.
Fibrosis, Laxity, and Mechanical Remodelling
In a collagen-variant auditory system, there may therefore be a spectrum between under-damped laxity and over-stiffened remodelling. Under-damped laxity may allow excessive vibration, pressure movement, resonance, or sensory intrusion. Over-stiffened remodelling may reduce fine mechanical responsiveness, alter acoustic impedance, or impair frequency tuning. Both states can distort sound transmission, but they may do so in different ways.
Fibrotic remodelling provides one route through which sensory tissues may become mechanically altered. Cochlear fibrosis research identifies pathways including TGF-β/SMAD, PDGF, and Wnt/β-catenin signalling as contributors to fibrotic change. Fibrosis and neo-ossification can increase acoustic impedance and worsen auditory outcomes. Although much of this literature arises from injury, implantation, or disease models, the broader mechanistic principle is relevant: excessive or disordered matrix deposition changes tissue mechanics.
This spectrum is important because auditory sensitivity is often assumed to reflect heightened perception or exaggerated reaction. A tissue-level framework suggests a more specific possibility: sound may be transmitted through an auditory apparatus with altered damping, stiffness, compliance, or pressure regulation. The downstream experience of sound intolerance may therefore reflect mechanical distortion as much as perceptual interpretation.
Pressure Dynamics and Reduced Containment
The structural boundaries of the ear also influence sensory experience. Superior semicircular canal dehiscence provides a useful example of how altered containment changes sound and vestibular processing. In this condition, thinning or opening of the temporal bone creates an abnormal third window within the inner ear. This redistributes pressure and vibration and may produce hyperacusis, autophony, tinnitus, vertigo, sound-induced dizziness, and heightened awareness of internal body sounds.
The relevance of this model extends beyond the specific diagnosis. It demonstrates that sound sensitivity can arise from altered biomechanics and pressure transmission rather than psychological intolerance. If containment is reduced, vibration and fluid movement are redistributed. The person is not reacting excessively to sound; the auditory and vestibular systems are physically transmitting sound and pressure differently.
Reports of superior semicircular canal dehiscence in people with Ehlers-Danlos syndrome are particularly relevant because they suggest that connective tissue fragility may contribute to over-transmission and sensory intrusion, not only to hearing loss. A collagen-divergent ear may be vulnerable to excessive internal sound, abnormal resonance, altered vestibular signalling, pressure sensitivity, and sound-induced disequilibrium.
Auditory Processing Downstream of Altered Input
Auditory processing 'disorder' is generally defined by difficulty interpreting sound despite hearing thresholds that may appear adequate on standard audiological testing. It may involve difficulty with speech-in-noise, localisation, sequencing, discrimination, rapid auditory analysis, temporal processing, and auditory memory. These difficulties are often treated as central processing problems, separate from the peripheral ear.
Auditory processing 'disorder' can affect the brain’s ability to separate, filter and prioritise sound. In environments with competing noise - such as cafés, classrooms, busy streets, or conversations where more than one person is speaking - speech may not stand out clearly from the background. Instead, the whole soundscape can arrive at once, making it difficult to isolate the voice that needs to be followed. This is not a hearing problem, because the sound itself may be heard; the difficulty lies in organising that sound into clear, usable meaning. For some people, this overlaps with ADHD attentional processing, where irrelevant sensory input cannot be easily filtered out and attention cannot be smoothly directed toward one auditory source. In this sense, auditory processing difficulties present as being unable to separate the one signal from the surrounding noise.
However, auditory processing does not occur independently of input quality; the brain processes the signal it receives. If that signal has already been altered by connective tissue variation, cochlear stiffness, poor damping, abnormal resonance, pressure instability, vestibular involvement, or matrix remodelling, then the cognitive burden of auditory processing is increased.
A person may therefore detect sound while still being overwhelmed by it. Speech may blur in noise not only because cortical sorting is difficult, but because the incoming acoustic stream is already unstable or crowded. Small sounds may feel disproportionately present because damping is reduced. Layered noise may become exhausting because the system receives too much undifferentiated acoustic detail. Certain frequencies may feel piercing because mechanical tuning or neural gain is altered. Internal sounds may be unusually prominent where pressure transmission is atypical. Auditory fatigue may accumulate because the system is working across several levels at once: tissue, transmission, vestibular regulation, neural gain, attention, memory, and meaning extraction.
Central gain models of hyperacusis remain relevant within this account, but they should not be treated as a complete explanation. Central amplification may occur downstream of a mechanically altered signal. In other words, the nervous system may amplify sound that has already been shaped by structurally divergent peripheral tissues.
From Sensory Structure to Behavioural Misclassification
The same pathway can be described across both vision and hearing: genomic variation may contribute to connective tissue divergence; connective tissue divergence may alter the peripheral sensory interface; altered sensory interfaces may change the quality of input; the nervous system receives information that is harsher, noisier, less filtered, less stable, or more demanding; neurological overload follows; the visible response is then interpreted through behavioural, psychiatric, or educational frameworks.
This produces a recurrent pattern of misclassification. Light sensitivity may be described as social withdrawal. Sound intolerance may be described as oppositional behaviour. Painful environments may be described as meltdowns. Auditory fatigue may be described as poor attention. Visual overload may be described as anxiety. Vestibular distress may be described as panic. Avoidance may be interpreted as refusal. The physiological burden is translated into behavioural language because the upstream tissue conditions have not been examined.
Behavioural descriptions can become clinically and educationally consequential. They influence how children are supported, how adults are treated, how environments are designed, and whether distress is accommodated or pathologised. Where sensory overload is rooted partly in connective tissue architecture, pressure dynamics, optical instability, auditory mechanics, inflammation, or tissue remodelling, purely behavioural interpretation risks obscuring the biological origin of the response.
Omission from Diagnostic Frameworks
The physiology described here is not entirely unknown. Collagen-related ocular and auditory phenotypes are recognised in syndromic connective tissue disorders. The extracellular matrix is known to influence sensory organ development and function. The tectorial membrane, basilar membrane, cornea, sclera, lens capsule, Bruch’s membrane, cochlear structures, and vestibular system are all dependent on precise tissue organisation. Epigenetic regulation is recognised in sensory development, injury response, and repair. Fibrosis is recognised as a mechanism of altered tissue mechanics.
The problem is not the absence of biological knowledge, but the failure of integration. These mechanisms are rarely brought into autism, ADHD, sensory processing, migraine, fatigue, hyperacusis, or auditory processing frameworks in a systematic way. Instead, sensory difficulty is commonly interpreted using binary diagnoses focused on central processing, psychology, behaviour, or psychiatric comorbidity. The sensory organs themselves are often treated as peripheral in the conceptual sense as well as the anatomical one.
A more adequate framework would require cross-disciplinary thinking and integration between genomics, ophthalmology, audiology, neurology, developmental biology, connective tissue medicine, immunology, epigenetics, and environmental health. It would also require recognition that divergent sensory experience may not be reducible to subjective intolerance. In some individuals, the body receiving the sensory environment may be structurally and mechanically different
Sensory overload in divergent populations should not be understood as a problem of perception, attention, behaviour, or central neurological filtering. The eye and ear are connective tissue-rich sensory interfaces whose structural properties shape visual and auditory input before it reaches the nervous system. Variation in collagen, fibrillin, extracellular matrix organisation, tissue compliance, pressure regulation, epigenetic expression, inflammatory repair, and fibrotic remodelling may therefore contribute to altered sensory experience at the level of signal formation.
Artist:Henry Gray (1918) Anatomy of the human body
Contact
Reach out with questions or collaboration ideas.
AChambers@divergentgenomics.org
© Alexandra Chambers 2026. All rights reserved.